Potenziale di membrana per il riposo e l'azione. Potenziale di riposo e potenziale d'azione
Potenziale di riposo
Le membrane, comprese le membrane plasmatiche, sono in linea di principio impermeabili alle particelle cariche. È vero, la membrana contiene Na + / K + -ATP-ase (Na + / K + -ATP-ase), che trasferisce attivamente ioni Na + dalla cellula in cambio di ioni K +. Questo trasporto è volatile ed è associato all'idrolisi dell'ATP (ATP). Grazie al lavoro della "pompa Na+, K+", viene mantenuta la distribuzione di non equilibrio degli ioni Na+ e K+ tra la cellula e l'ambiente. Poiché la scissione di una molecola di ATP assicura il trasferimento di tre ioni Na + (dalla cellula) e due ioni K + (alla cellula), questo trasporto è elettrogeno, cioè . il citoplasma della cellula è caricato negativamente rispetto allo spazio extracellulare.
Potenziale elettrochimico. Il contenuto delle celle è caricato negativamente rispetto allo spazio extracellulare. La ragione principale per la comparsa sulla membrana di un potenziale elettrico (potenziale di membrana Δψ) è l'esistenza canali ionici specifici. Il trasporto di ioni attraverso i canali avviene lungo un gradiente di concentrazione o sotto l'azione di un potenziale di membrana. In una cellula non eccitata, parte dei canali K + è allo stato aperto e gli ioni K + diffondono costantemente dal neurone nell'ambiente (lungo il gradiente di concentrazione). All'uscita dalla cella, gli ioni K+ portano via una carica positiva, che crea un potenziale di riposo pari a circa -60 mV. Si può vedere dai coefficienti di permeabilità dei vari ioni che i canali permeabili a Na+ e Cl- sono prevalentemente chiusi. Gli ioni fosfato e gli anioni organici come le proteine non sono praticamente in grado di passare attraverso le membrane. Utilizzando l'equazione di Nernst (RT / ZF, dove R è la costante del gas, T è la temperatura assoluta, Z è la valenza ionica, F è il numero di Faraday), si può dimostrare che il potenziale di membrana di una cellula nervosa è principalmente determinato da ioni K +, che danno il contributo principale alla conduttività della membrana.
Canali ionici... Nelle membrane della cellula nervosa sono presenti canali permeabili per gli ioni Na+, K+, Ca2+ e Cl-. Questi canali sono spesso chiusi e aperti solo per un breve periodo. I canali sono suddivisi in voltaggio-dipendenti (o elettroeccitabili), come i canali veloci Na+, e ligando-dipendenti (o chimicamente eccitabili), come i recettori colinergici nicotinici. I canali sono proteine integrali di membrana composte da molte subunità. A seconda del cambiamento nel potenziale di membrana o dell'interazione con i corrispondenti ligandi, neurotrasmettitori e neuromodulatori (vedi Fig. 343), le proteine recettrici possono trovarsi in uno dei due stati conformazionali, che determina la permeabilità del canale ("aperto" - "chiuso" - ed ecc.).
Trasporto attivo:
La stabilità del gradiente ionico si ottiene attraverso il trasporto attivo: le proteine di membrana trasportano gli ioni attraverso la membrana contro gradienti elettrici e/o di concentrazione, consumando per questo energia metabolica. Il processo di trasporto attivo più importante è il funzionamento della pompa Na/K, presente in quasi tutte le cellule; la pompa pompa ioni sodio fuori dalla cellula mentre contemporaneamente pompa ioni potassio nella cellula. Viene così assicurata una bassa concentrazione intracellulare di ioni sodio e un'alta concentrazione di ioni potassio. Il gradiente di concentrazione degli ioni sodio sulla membrana ha funzioni specifiche associate alla trasmissione di informazioni sotto forma di impulsi elettrici, nonché al mantenimento di altri meccanismi di trasporto attivo e regolazione del volume cellulare. Pertanto, non sorprende che più di 1/3 dell'energia consumata dalla cella venga spesa per la pompa Na / K e, in alcune delle celle più attive, fino al 70% dell'energia venga spesa per il suo funzionamento .
Trasporto passivo:
I processi di diffusione e trasporto liberi forniti dai canali ionici e dai vettori sono effettuati lungo un gradiente di concentrazione o un gradiente di carica elettrica (collettivamente indicato come gradiente elettrochimico). Tali meccanismi di trasporto sono classificati come "trasporto passivo". Ad esempio, secondo questo meccanismo, il glucosio dal sangue entra nelle cellule, dove la sua concentrazione è molto più alta.
Pompa ionica:
Le pompe ioniche (pompe) sono proteine integrali che forniscono il trasporto attivo di ioni contro il gradiente di concentrazione. L'energia per il trasporto è l'energia dell'idrolisi dell'ATP. Distinguere tra pompa Na + / K + (pompa Na + dalla cella in cambio di K +), pompa Ca ++ (pompa Ca ++ dalla cella), pompa Cl– (pompa fuori Cl - dalla cella) .
Come risultato del funzionamento delle pompe ioniche, vengono creati e mantenuti gradienti ionici transmembrana:
La concentrazione di Na +, Ca ++, Cl - all'interno della cellula è inferiore a quella esterna (nel fluido intercellulare);
La concentrazione di K + all'interno della cellula è superiore a quella esterna.
Pompa sodio - potassio- è una proteina speciale che permea l'intero spessore della membrana, che pompa costantemente ioni di potassio nella cellula, mentre contemporaneamente pompa fuori di essa ioni di sodio; in questo caso, il movimento di entrambi gli ioni avviene contro i gradienti delle loro concentrazioni. Queste funzioni sono possibili grazie a due importanti proprietà di questa proteina. Innanzitutto, la forma della molecola portatrice può cambiare. Questi cambiamenti si verificano a seguito dell'aggiunta di un gruppo fosfato alla molecola di trasporto a causa dell'energia rilasciata durante l'idrolisi dell'ATP (cioè la decomposizione dell'ATP in ADP e il resto dell'acido fosforico). In secondo luogo, questa stessa proteina agisce come ATP-asi (cioè un enzima che idrolizza l'ATP). Poiché questa proteina svolge il trasporto di sodio e potassio e, inoltre, ha attività ATP-asica, viene chiamata “ATP-asi sodio-potassio”.
Semplificando, l'azione della pompa sodio-potassio può essere rappresentata come segue.
1. Dal lato interno della membrana, gli ioni ATP e sodio entrano nella molecola proteica di trasporto e dal lato esterno, gli ioni potassio.
2. La molecola vettore idrolizza una molecola di ATP.
3. Con la partecipazione di tre ioni sodio dovuta all'energia dell'ATP, il resto dell'acido fosforico viene aggiunto al vettore (fosforilazione del vettore); anche questi tre ioni sodio si legano al trasportatore.
4. Come risultato dell'aggiunta del residuo di acido fosforico, si verifica un tale cambiamento nella forma della molecola portatrice (conformazione) che gli ioni sodio compaiono dall'altra parte della membrana, già all'esterno della cellula.
5. Tre ioni sodio vengono rilasciati nell'ambiente esterno e, al loro posto, due ioni potassio vengono combinati con il vettore fosforilato.
6. L'aggiunta di due ioni potassio provoca la defosforilazione del vettore - il rilascio del resto di acido fosforico ad essi.
7. La defosforilazione, a sua volta, provoca una tale conformazione del trasportatore che gli ioni potassio compaiono dall'altra parte della membrana, all'interno della cellula.
8. Gli ioni potassio vengono rilasciati all'interno della cellula e l'intero processo viene ripetuto.
L'importanza della pompa sodio-potassio per la vita di ogni cellula e dell'organismo nel suo insieme è determinata dal fatto che il pompaggio continuo di sodio dalla cellula e l'iniezione di potassio in essa è necessario per l'attuazione di molte funzioni vitali processi: osmoregolazione e conservazione del volume cellulare, mantenimento della differenza di potenziale su entrambi i lati della membrana, mantenimento dell'attività elettrica nelle cellule nervose e muscolari, per il trasporto attivo attraverso le membrane di altre sostanze (zuccheri, amminoacidi). Grandi quantità di potassio sono necessarie anche per la sintesi proteica, la glicolisi, la fotosintesi e altri processi. Circa un terzo di tutto l'ATP consumato da una cellula animale a riposo viene speso proprio per il mantenimento della pompa sodio-potassio. Se per qualche influenza esterna la respirazione della cellula viene soppressa, cioè l'apporto di ossigeno e la produzione di ATP viene interrotta, allora la composizione ionica del contenuto interno della cellula inizierà a cambiare gradualmente. Alla fine entrerà in equilibrio con la composizione ionica dell'ambiente che circonda la cellula; in questo caso si verifica la morte.
Potenziale d'azione di una cellula eccitabile e sua fase:
P.D, - rapida fluttuazione del potenziale di membrana derivante dall'eccitazione dei nervi., Mouse. E altre cellule possono diffondersi.
1. fase di salita
2.reversion o overshoot (carica dei flip)
3. Ripristino della polarità o ripolarizzazione
4. potenziale di traccia positivo
5. Impronta negativa. Potenziale
Risposta locale-è il processo di risposta della membrana a uno stimolo in un'area specifica del neurone. Non diffuso sugli assoni. Maggiore è lo stimolo, più cambia la risposta locale. Allo stesso tempo, il livello di depolarizzazione non raggiunge quello critico, rimane sottosoglia. Di conseguenza, una risposta locale può esercitare effetti elettrotonici sulle regioni di membrana adiacenti, ma non può propagarsi allo stesso modo di un potenziale d'azione. L'eccitabilità della membrana in luoghi di depolarizzazione locale e in luoghi di depolarizzazione elettrotonica causata da essa è aumentata.

Attivazione e inattivazione del sistema del sodio:
L'impulso di corrente depolarizzante porta all'attivazione dei canali del sodio e ad un aumento della corrente del sodio. Ciò fornisce una risposta locale. Lo spostamento del potenziale di membrana a un livello critico porta a una rapida depolarizzazione della membrana cellulare e fornisce un fronte per la crescita del potenziale d'azione. Se rimuovi lo ione Na + dall'ambiente esterno, il potenziale d'azione non sorge. Un effetto simile è stato ottenuto aggiungendo alla soluzione di perfusione TTX (tetrodotossina), uno specifico bloccante dei canali del sodio. Quando si utilizza il metodo "morsetto di tensione", è stato dimostrato che in risposta all'azione della corrente depolarizzante, una corrente in ingresso a breve termine (1-2 ms) scorre attraverso la membrana, che viene sostituita dopo un po' dalla corrente in uscita corrente (Fig. 2.11). Quando si sostituiscono gli ioni di sodio con altri ioni e sostanze, ad esempio la colina, è stato possibile dimostrare che la corrente in ingresso è fornita da una corrente di sodio, cioè, in risposta a uno stimolo depolarizzante, si verifica un aumento della conduttività del sodio (gNa +). Pertanto, lo sviluppo della fase di depolarizzazione del potenziale d'azione è dovuto ad un aumento della conduttività del sodio.

Consideriamo il principio di funzionamento dei canali ionici usando l'esempio di un canale del sodio. Si ritiene che il canale del sodio sia chiuso a riposo. Quando la membrana cellulare viene depolarizzata a un certo livello, le porte di attivazione m vengono aperte (attivazione) e il flusso di ioni Na + nella cellula aumenta. Pochi millisecondi dopo l'apertura della m-gate, la n-gate situata all'uscita dei canali del sodio si chiude (inattivazione) (Fig. 2.4). L'inattivazione si sviluppa molto rapidamente nella membrana cellulare e il grado di inattivazione dipende dall'entità e dalla durata dello stimolo depolarizzante.
Il lavoro dei canali del sodio è determinato dal valore del potenziale di membrana secondo determinate leggi di probabilità. Si calcola che il canale del sodio attivato lascia passare solo 6000 ioni in 1 ms. In questo caso, la corrente di sodio molto significativa che attraversa le membrane durante l'eccitazione è la somma di migliaia di singole correnti.
Quando un singolo potenziale d'azione viene generato in una spessa fibra nervosa, il cambiamento nella concentrazione di ioni Na + nell'ambiente interno è solo 1/100000 del contenuto interno di ioni Na nell'assone di calamaro gigante. Tuttavia, per le fibre nervose sottili, questo cambiamento di concentrazione può essere piuttosto significativo.
Oltre al sodio, nelle membrane cellulari sono installati altri tipi di canali, selettivamente permeabili ai singoli ioni: K+, Ca2+, e per questi ioni esistono varietà di canali (vedi Tabella 2.1).
Hodgkin e Huxley hanno formulato il principio di "indipendenza" dei canali, secondo il quale i flussi di sodio e potassio attraverso la membrana sono indipendenti l'uno dall'altro.
Cambiamento nell'eccitabilità quando eccitato:
1. Refrattarietà assoluta - ad es. completa ineccitabilità, determinata prima dal pieno impiego del meccanismo del "sodio", e poi dall'inattivazione dei canali del sodio (questo corrisponde grosso modo al picco del potenziale d'azione).
2. Refrattarietà relativa - es. ridotta eccitabilità associata alla parziale inattivazione del sodio e allo sviluppo dell'attivazione del potassio. In questo caso, la soglia viene aumentata e la risposta [AP] viene abbassata.
3. Esaltazione - es. l'ipereccitabilità è una supernormalità derivante dalla depolarizzazione in tracce.
4. Subnormalità - cioè ridotta eccitabilità derivante dall'iperpolarizzazione della traccia. Le ampiezze del potenziale d'azione nella fase di negatività della traccia sono in qualche modo ridotte e, sullo sfondo della positività della traccia, sono leggermente aumentate.
La presenza di fasi refrattarie determina la natura intermittente (discreta) della segnalazione nervosa e il meccanismo ionico del potenziale d'azione assicura la standardizzazione del potenziale d'azione (impulsi nervosi). In questa situazione, i cambiamenti nei segnali esterni sono codificati solo da un cambiamento nella frequenza del potenziale d'azione (codice di frequenza) o da un cambiamento nel numero di potenziali d'azione.

Per condurre un segnale da una cellula precedente alla successiva, il neurone genera segnali elettrici al suo interno. I movimenti dei tuoi occhi durante la lettura di questo paragrafo, la sensazione di una sedia morbida sotto il tuo sedere, la percezione della musica dalle cuffie e molto altro si basano sul fatto che centinaia di miliardi di segnali elettrici ti attraversano. Tale segnale può avere origine nel midollo spinale e viaggiare lungo un lungo assone fino alla punta del dito del piede. Oppure può coprire una distanza trascurabile nelle profondità del cervello, confinandosi ai limiti di un interneurone con processi brevi. Qualsiasi neurone che riceve un segnale lo invia attraverso il suo corpo e le sue escrescenze, e questo segnale è di natura elettrica.
Nel 1859, gli scienziati sono stati in grado di misurare la velocità con cui vengono trasmessi questi segnali elettrici. Si è scoperto che l'elettricità trasmessa da un assone vivente è fondamentalmente diversa dalla corrente elettrica nei metalli. Attraverso un filo metallico viene trasmesso un segnale elettrico ad una velocità prossima a quella della luce (300.000 chilometri al secondo), perché nel metallo sono presenti molti elettroni liberi. Tuttavia, nonostante questa velocità, il segnale è notevolmente indebolito, superando lunghe distanze. Se i segnali fossero trasmessi lungo gli assoni nello stesso modo in cui vengono trasmessi nei metalli, l'impulso nervoso proveniente dalle terminazioni nervose nella pelle dell'alluce decadrebbe completamente prima di raggiungere il cervello - la resistenza elettrica della materia organica è troppo alta , e il segnale è troppo debole...
Gli studi hanno dimostrato che l'elettricità viaggia molto più lentamente lungo gli assoni rispetto ai fili e che questa trasmissione si basa su un meccanismo precedentemente sconosciuto che fa sì che il segnale viaggi a una velocità di circa 30 metri al secondo. I segnali elettrici che viaggiano lungo i nervi, a differenza dei segnali che viaggiano attraverso i fili, non si indeboliscono mentre viaggiano. La ragione di ciò è che le terminazioni nervose non passano il segnale attraverso se stesse passivamente, semplicemente permettendo alle particelle cariche in esse di trasmetterlo l'una all'altra. In ogni punto sono un emettitore attivo di questo segnale, lo ritrasmettono, e una descrizione dettagliata di questo meccanismo richiederà un capitolo separato. Quindi, sacrificando l'alta velocità degli impulsi nervosi, a causa della trasmissione attiva del segnale, il neurone riceve la garanzia che il segnale che è sorto nell'alluce raggiungerà il midollo spinale senza indebolirsi affatto.

Per osservare il passaggio di un'onda di eccitazione elettrica, o potenziale d'azione (potenziale d'azione ['ækʃən pə'tenʃəl]), in una cellula vivente è sufficiente un semplice dispositivo: un'estremità di un sottile filo metallico è posta sulla superficie esterna dell'assone di un neurone sensoriale nella pelle, e l'altra è portata a un registratore che traccia una linea verso l'alto quando il segnale è amplificato e verso il basso quando il segnale è indebolito. Ogni contatto con la pelle innesca uno o più potenziali d'azione. Ad ogni potenziale, il registratore disegna un picco lungo e stretto.
Il potenziale d'azione di un neurone sensoriale dura solo circa 0,001 secondi e comprende due fasi: un rapido aumento, raggiungendo un picco, e poi un declino quasi altrettanto rapido dell'eccitazione, che porta alla posizione iniziale. E qui il registratore riporta un fatto inaspettato: tutti i potenziali d'azione che sorgono in una stessa cellula nervosa sono approssimativamente gli stessi. Questo può essere visto nell'immagine a sinistra: tutti i picchi disegnati dal registratore hanno approssimativamente la stessa forma e ampiezza, non importa quanto duro o quanto sia stato il tocco sulla pelle che li ha causati. Un leggero colpo o un pizzico tangibile saranno trasmessi da potenziali d'azione della stessa grandezza. Il potenziale d'azione è un segnale costante che obbedisce al principio del "tutto o niente": dopo che lo stimolo supera un certo valore di soglia, appare sempre approssimativamente lo stesso segnale, né più né meno del solito. E se lo stimolo è inferiore al valore di soglia, il segnale non verrà trasmesso affatto: ad esempio, puoi toccare la pelle così facilmente con la punta della penna che questo tocco non si farà sentire.
Il principio del tutto o niente nell'emergere di un potenziale d'azione solleva nuove domande. In che modo il neurone sensoriale segnala la forza dello stimolo: pressione forte o debole, luce brillante o fioca? Come riporta la durata dello stimolo? Infine, come fanno i neuroni a distinguere un tipo di informazione sensoriale da un altro - per esempio, come distinguono il tatto dal dolore, la luce, l'odore o il suono? E come distinguono le informazioni sensoriali per la percezione dalle informazioni motorie per l'azione?

L'evoluzione ha risolto il problema di come comunicare la forza di uno stimolo utilizzando lo stesso tipo di segnali della stessa grandezza: questa forza è determinata frequenza(frequenza ['friːkwənsɪ]) con cui vengono emessi potenziali d'azione. Uno stimolo debole, come un tocco leggero sul braccio, emette solo due o tre potenziali d'azione al secondo, mentre una forte pressione, come pizzicare o colpire il gomito, può causare un'esplosione di centinaia di potenziali d'azione al secondo. In questo caso, la durata della sensazione è determinata dalla durata dell'emergere dei potenziali d'azione.
I neuroni usano codici elettrici diversi per dire al cervello che trasportano informazioni su stimoli diversi, come dolore, luce o suono? Si è scoperto che no! È sorprendente, ma c'è pochissima differenza tra i potenziali d'azione generati da neuroni di diversi sistemi sensoriali (ad esempio, visivo o tattile)! Pertanto, il carattere e la natura della sensazione non dipendono dalle differenze nei potenziali d'azione (il che apre una prospettiva piuttosto eccitante per pensare alla "matrice" del film omonimo). Un neurone che trasmette informazioni uditive è progettato esattamente allo stesso modo di un neurone proveniente dal circuito nervoso visivo e conducono gli stessi potenziali d'azione allo stesso modo. Senza sapere a quale catena nervosa appartiene un particolare neurone, è impossibile determinare quali informazioni trasporta solo analizzando il suo funzionamento.
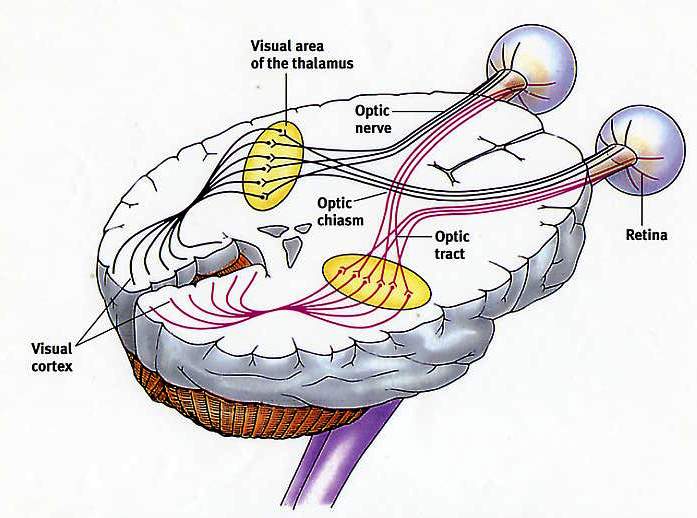
La natura delle informazioni trasmesse dipende principalmente dal tipo di fibre nervose eccitate e dai sistemi cerebrali specifici con cui queste fibre sono collegate. Le sensazioni di ogni tipo vengono trasmesse lungo i propri percorsi e il tipo di informazione trasmessa da un neurone dipende proprio dal percorso di cui questo neurone fa parte. In qualsiasi percorso sensoriale, le informazioni viaggiano dal primo neurone sensoriale (un recettore che risponde a uno stimolo esterno, come il tatto, l'olfatto o la luce) a neuroni specializzati nel midollo spinale o nel cervello. Pertanto, le informazioni visive differiscono dalle informazioni uditive solo in quanto vengono trasmesse lungo altri percorsi, a partire dalla retina dell'occhio e terminando nella parte del cervello responsabile della percezione visiva.
Anche i segnali inviati dai motoneuroni nel cervello ai muscoli sono quasi identici a quelli inviati dai neuroni sensoriali dalla pelle al cervello. Obbediscono allo stesso principio del "tutto o niente", trasmettono anche l'intensità del segnale usando la frequenza dei potenziali d'azione, e anche il risultato del segnale dipende solo da quale circuito nervoso è incluso questo neurone. Quindi, una rapida successione di potenziali d'azione, seguendo un certo percorso conduttivo, provoca proprio il movimento delle dita, e non, diciamo, la percezione di luci colorate, solo perché questo percorso è associato ai muscoli delle mani, e non la retina degli occhi.

L'universalità dei potenziali d'azione non si limita alla somiglianza della loro manifestazione in diversi neuroni situati all'interno dello stesso organismo. Sono così simili in animali diversi che anche un ricercatore esperto non è in grado di distinguere con precisione la registrazione del potenziale d'azione della fibra nervosa di una balena, un topo, una scimmia o il suo consulente scientifico. Tuttavia, i potenziali d'azione in cellule diverse non sono identici: c'è ancora una leggera differenza nella loro ampiezza e durata, e l'affermazione "tutti i potenziali d'azione sono uguali" è altrettanto imprecisa quanto "tutte le buganvillee sono uguali".
Quindi, ogni neurone trasmette un segnale attraverso il suo corpo ed elabora allo stesso modo. Tutta la varietà di informazioni che riceviamo dai neuroni sensoriali, tutti i movimenti che il nostro corpo può compiere sono il risultato della trasmissione di un unico tipo di segnale all'interno dei neuroni. Rimane una "sciocchezza": capire che tipo di segnale è e come viene trasmesso.

Separiamo abitualmente tutto ciò che consideriamo natura vivente, compresi noi stessi, dalle cose "inanimate", compresi i metalli e la corrente elettrica trasmessa attraverso di essi. È tanto più sorprendente rendersi conto che i metalli non sono solo presenti nel nostro corpo: sono necessari, senza di loro il corpo non può esistere. La corrente elettrica non è un fenomeno una tantum, ma si verifica continuamente in centinaia di miliardi di neuroni, che sono penetrati in tutto il nostro corpo con i loro processi. In questo momento, puoi percepire ogni sorta di segno della sua presenza: il fatto che tu sia a conoscenza di questo testo è il risultato di innumerevoli trasmissioni di corrente elettrica. La sensazione di fame e piacere dall'odore del cibo cucinato, la percezione stessa di questo odore, il tocco del vento che soffia attraverso la finestra sulla tua pelle ... L'elenco è infinito. E il desiderio di capire come tutto ciò avvenga consiste anche negli impulsi elettrici che si originano nei neuroni.
Poiché lo scopo di questo capitolo è quello di comunicare solo le informazioni più generali sul passaggio di un impulso nervoso, qui è necessario considerare l'ambiente in cui sorge, quelle condizioni nella cellula che ne rendono possibile l'insorgenza e la trasmissione. Vale quindi la pena iniziare studiando la testa di ponte su cui si svilupperanno gli eventi, ovvero dal neurone in stato di riposo (stato dormiente [‘dɔːmənt steɪt]).

A metà del secolo scorso, gli scienziati hanno trovato un modo per stabilire in quale parte del neurone è presente una carica elettrica. Per farlo, usa voltmetro (voltmetro ['vəultˌmiːtə]) (dispositivo per la misura della tensione di campo elettrico) con due elettrodi. Un elettrodo è posto all'interno del neurone, posizionandolo vicino alla membrana cellulare, e il secondo elettrodo si trova nell'ambiente che circonda il neurone, dall'altra parte della stessa membrana. Il voltmetro mostra che su diversi lati della membrana cellulare ci sono cariche elettriche, negativo all'interno della cellula e positivo all'esterno. L'esistenza di tali cariche elettriche di polarità opposta su entrambi i lati della membrana crea un campo elettrico, una caratteristica importante della quale è potenziale... Il potenziale, in termini semplici, è la capacità di svolgere un lavoro, ad esempio il lavoro di trascinare una particella carica da un luogo all'altro. Più cariche negative si sono accumulate su un lato e più cariche positive sull'altro lato della membrana, più forte è il campo elettrico che creano e più forza sono in grado di trascinare avanti e indietro le particelle cariche. La differenza tra cariche elettriche esterne e interne è chiamata potenziale di membrana (potenziale di membrana ['membreɪn pə'tenʃəl]) riposo. Per un neurone, è circa 70 mV (millivolt), cioè 70 millesimi di volt o sette centesimi di volt. Per fare un confronto, la differenza di potenziale in una batteria AA è di 1,5 volt - 20 volte di più. Cioè, il potenziale di membrana a riposo di un neurone è solo 20 volte più debole di quello tra i terminali di una batteria AA - piuttosto grande, a quanto pare. Il potenziale elettrico esiste solo sulla membrana e nelle altre sue parti il neurone è elettricamente neutro.
Più precisamente, il potenziale di membrana a riposo di un neurone è -70 mV (meno settanta millivolt). Il segno meno significa solo che la carica negativa è all'interno della cellula, e non all'esterno, e quindi il campo elettrico generato è in grado di trascinare ioni caricati positivamente attraverso la membrana nella cellula.
Attori nella creazione del potenziale di membrana a riposo:

1 ... V membrana cellulare un neurone, ci sono canali attraverso i quali gli ioni che trasportano una carica elettrica possono attraversarlo. Allo stesso tempo, la membrana non è solo una "partizione" passiva tra l'ambiente interno del neurone e il fluido intercellulare che lo circonda: speciali proteine incastonate nella carne della membrana aprono e chiudono questi canali, e quindi la membrana controlla il passaggio di ioni - atomi con una carica elettrica. Accumulando ioni caricati negativamente all'interno della cellula, il neurone aumenta il numero di cariche negative all'interno, portando così ad un aumento delle cariche positive all'esterno, e quindi il potenziale elettrico aumenta. Poiché un protone ha una carica positiva e un elettrone è negativo, quindi con un eccesso di protoni si ottiene uno ione con carica positiva e con un eccesso di elettroni, uno con carica negativa. Se desideri maggiori informazioni su atomi e ioni, puoi tornare a. È importante capire che il potenziale di membrana esiste proprio al confine della membrana cellulare e che i fluidi in generale all'interno e all'esterno del neurone rimangono elettricamente neutri. Gli ioni, per i quali la membrana è permeabile, rimangono vicino ad essa, poiché le cariche positive e negative sono reciprocamente attratte l'una dall'altra. Di conseguenza, uno strato di ioni positivi "seduto" su di esso si forma all'esterno della membrana e ioni negativi all'interno. Pertanto, la membrana svolge il ruolo di una capacità elettrica che separa le cariche, all'interno della quale è presente un campo elettrico. La membrana è quindi un condensatore naturale.
2 . proteine con carica negativa situato all'interno del neurone vicino alla superficie interna della membrana. La carica delle proteine rimane sempre la stessa ed è solo una parte della carica totale della superficie interna della membrana. A differenza degli ioni, le proteine non possono lasciare la cellula ed entrarvi - sono troppo grandi per questo. La carica totale cambia a seconda del numero di ioni caricati positivamente vicino alla membrana, la cui concentrazione può cambiare a causa della loro transizione dalla cellula all'esterno e dall'esterno all'interno.
3 ... ioni di potassio caricati positivamente (K +) possono muoversi liberamente tra l'ambiente interno ed esterno quando il neurone è a riposo. Si muovono attraverso costantemente aperti canali di flusso del potassio (flusso di potassio passaggio), attraverso il quale possono passare solo gli ioni K + e nient'altro. I canali di flusso sono chiamati canali che non hanno porte, il che significa che sono aperti in qualsiasi stato del neurone. Ci sono molti più ioni potassio all'interno della cellula che all'esterno. Ciò è dovuto al funzionamento costante della pompa sodio-potassio (sarà discusso di seguito), quindi, nello stato di riposo del neurone, gli ioni K + iniziano a spostarsi nell'ambiente esterno, poiché la concentrazione della stessa sostanza tende a pareggiare nel sistema generale. Se versiamo una sostanza in una piscina con acqua in un angolo, la sua concentrazione in questo angolo sarà molto alta e in altre parti della piscina - zero o molto piccola. Tuttavia, dopo un po' scopriremo che la concentrazione di questa sostanza si è stabilizzata in tutto il bacino a causa del moto browniano. In questo caso parlano della "pressione parziale" di una determinata sostanza, sia essa un liquido o un gas. Se l'alcol viene versato in un angolo della piscina, ci sarà una grande differenza nella concentrazione di alcol tra quell'angolo e il resto della piscina. Ci sarà una pressione parziale delle molecole di alcol e gradualmente saranno distribuite uniformemente sul bacino in modo che la pressione parziale scompaia, poiché la concentrazione delle molecole di alcol ovunque si equalizzerà. Pertanto, gli ioni K + portano via la carica positiva dal neurone, lasciando fuori a causa della pressione parziale, che è più forte della forza attrattiva delle proteine caricate negativamente, se la differenza nella concentrazione di ioni all'interno e all'esterno della cellula è abbastanza grande . Poiché le proteine con carica negativa rimangono all'interno, si forma così una carica negativa sul lato interno della membrana. Per una chiara comprensione del lavoro dei meccanismi cellulari, è importante ricordare che, nonostante il costante deflusso di ioni potassio dalla cellula, ce ne sono sempre più all'interno del neurone che all'esterno.
4 ... ioni sodio caricati positivamente (Na +) si trovano all'esterno della membrana e creano lì una carica positiva. Durante la fase di riposo del neurone, i canali del sodio della cellula Chiuso, e Na + non possono passare all'interno e la loro concentrazione all'esterno aumenta a causa del lavoro della pompa sodio-potassio, che li rimuove dal neurone.
5 ... il ruolo di carica negativa ioni cloro (Cl -) e carica positivamente ioni calcio (Ca 2+) creare un potenziale di membrana è piccolo, quindi il loro comportamento rimarrà dietro le quinte.
Formazione del potenziale di membrana a riposo avviene in due fasi:
Fase I... una piccola differenza di potenziale (-10 mV) viene creata usando pompa sodio potassio.

A differenza di altri canali della membrana, il canale sodio-potassio è in grado di far passare attraverso se stesso sia ioni sodio che potassio. Inoltre, Na + può attraversarlo solo dalla cellula verso l'esterno e K + dall'esterno verso l'interno. Un ciclo di funzionamento di questo canale comprende 4 fasi:
1 ... La "porta" del canale sodio-potassio è aperta solo dal lato interno della membrana e 3 Na + entrano lì

2 ... la presenza di Na + all'interno del canale lo influenza in modo che possa distruggere parzialmente una molecola ATF(ATP) ( adenosina trifosfato), (trifosfato di adenosina [ə'dɛnəsiːn trai'fɔsfeɪt]) che è la "batteria" della cellula, che immagazzina energia e la cede quando necessario. Con una tale distruzione parziale, che consiste nell'eliminazione di un gruppo fosfato PO 4 3− dall'estremità della molecola, viene rilasciata energia, che viene esattamente spesa per il trasferimento di Na + nello spazio esterno.
3 ... quando il canale si apre in modo che Na + esca, rimane aperto e due ioni K + vi entrano - sono attratti da cariche negative di proteine dall'interno. Il fatto che solo due ioni potassio siano posti nel canale contenente tre ioni sodio è abbastanza logico: l'atomo di potassio ha un diametro maggiore.
4 ... la presenza di ioni potassio ora, a sua volta, influenza il canale in modo che le "porte" esterne siano chiuse e quelle interne si aprano e K + entri nell'ambiente interno del neurone.
Ecco come funziona la pompa sodio-potassio, "scambiando" tre ioni sodio con due ioni potassio. Poiché la carica elettrica di Na + e K + è la stessa, si scopre che tre cariche positive vengono rimosse dalla cellula e solo due entrano. A causa di ciò, la carica positiva interna della membrana cellulare diminuisce e quella esterna aumenta. Inoltre, si crea una differenza nella concentrazione di Na + e K + su diversi lati della membrana:
=) ci sono molti ioni sodio all'esterno della cellula e pochi all'interno. Allo stesso tempo, i canali del sodio sono chiusi e Na + non può tornare nella cellula e non si allontana dalla membrana, poiché è attratto dalla carica negativa esistente sul lato interno della membrana.
=) ci sono molti ioni potassio all'interno della cellula, ma ce ne sono pochi all'esterno, e questo porta al deflusso di K+ dalla cellula attraverso i canali del potassio che si aprono durante la fase di riposo del neurone.

Fase II la formazione del potenziale di membrana a riposo si basa proprio su questo deflusso di ioni potassio dal neurone. La figura a sinistra mostra la composizione ionica della membrana all'inizio del secondo stadio della formazione del potenziale di riposo: molto K + e proteine caricate negativamente (designate A 4-) all'interno, e Na + attaccato al membrana esterna. Spostandosi nell'ambiente esterno, gli ioni potassio portano via le loro cariche positive dalla cellula, mentre la carica totale della membrana interna diminuisce. Proprio come gli ioni sodio positivi, gli ioni potassio fuoriusciti dalla cellula rimangono all'esterno della membrana, attratti dalla carica negativa interna, e la carica positiva esterna della membrana è la somma delle cariche Na+ e K+. Nonostante la perdita attraverso i canali di flusso, ci sono sempre più ioni potassio all'interno della cellula che all'esterno.
Sorge la domanda: perché gli ioni potassio non continuano a defluire fino al momento in cui il loro numero all'interno della cellula e all'esterno diventa lo stesso, cioè fino a quando la pressione parziale creata da questi ioni scompare? La ragione di ciò è che quando il K + lascia la cella, la carica positiva aumenta all'esterno e all'interno si forma un eccesso di carica negativa. Ciò riduce il desiderio che gli ioni potassio lascino la cellula, perché la carica positiva esterna li respinge e quella negativa interna li attrae. Pertanto, dopo qualche tempo, K + smette di defluire nonostante il fatto che nell'ambiente esterno la loro concentrazione sia inferiore a quella interna: l'influenza delle cariche su diversi lati della membrana supera la forza della pressione parziale, cioè supera la tendenza del K+ a distribuirsi uniformemente nel liquido all'interno e all'esterno del neurone. Al momento del raggiungimento di questo equilibrio, il potenziale di membrana del neurone si ferma a circa -70 mV.
Non appena il neurone raggiunge il potenziale di membrana a riposo, è pronto per l'emergenza e la conduzione del potenziale d'azione, di cui parleremo nel prossimo capitolo citologico.
Quindi, per riassumere: la distribuzione irregolare degli ioni potassio e sodio su entrambi i lati della membrana è causata dall'azione di due forze in competizione: a) la forza di attrazione e repulsione elettrica, eb) la forza di pressione parziale derivante da una differenza di concentrazione. Il lavoro di queste due forze rivali si svolge nelle condizioni dell'esistenza di canali sodio, potassio e sodio-potassio diversamente disposti, che agiscono come regolatori dell'azione di queste forze. Il canale del potassio è un canale di flusso, cioè è sempre aperto a riposo del neurone, in modo che gli ioni K+ possano facilmente camminare avanti e indietro sotto l'influenza delle forze di repulsione/attrazione elettrica e sotto l'influenza di forze causato dalla pressione parziale, cioè dalla differenza nella concentrazione di questi ioni. Il canale del sodio è sempre chiuso a riposo del neurone, per cui gli ioni Na+ non possono attraversarlo. E infine, il canale sodio-potassio, progettato in modo tale da funzionare come una pompa, che spinge tre ioni sodio verso l'esterno ad ogni ciclo e spinge due ioni potassio verso l'interno.
Tutta questa costruzione garantisce l'emergere del potenziale di membrana a riposo del neurone: ad es. uno stato in cui si realizzano due cose:
a) c'è una carica negativa all'interno e una carica positiva all'esterno.
b) ci sono molti ioni K + all'interno, attaccati alle parti caricate negativamente delle proteine, e quindi si verifica una pressione parziale di potassio - la tendenza degli ioni di potassio ad uscire per equalizzare la concentrazione.
c) ci sono molti ioni Na+ all'esterno, che formano in parte coppie con ioni Cl-. E quindi c'è una pressione parziale di sodio - il desiderio degli ioni sodio di entrare nella cellula per equalizzare la concentrazione.
Come risultato del funzionamento della pompa potassio-sodio, si ottengono tre forze esistenti sulla membrana: l'intensità del campo elettrico e l'intensità di due pressioni parziali. Queste forze iniziano a funzionare quando il neurone lascia lo stato di riposo.
Ogni cellula vivente è ricoperta da una membrana semipermeabile attraverso la quale avvengono il movimento passivo e il trasporto selettivo attivo di ioni carichi positivamente e negativamente. A causa di questo trasferimento, c'è una differenza nelle cariche elettriche (potenziali) tra le superfici esterne ed interne della membrana: il potenziale di membrana. Ci sono tre manifestazioni distinte del potenziale di membrana: potenziale di membrana a riposo, potenziale locale, o risposta locale, e potenziale d'azione.
Se gli stimoli esterni non agiscono sulla cellula, il potenziale di membrana rimane costante per lungo tempo. Il potenziale di membrana di una tale cellula a riposo è chiamato potenziale di membrana a riposo. Per la superficie esterna della membrana cellulare, il potenziale di riposo è sempre positivo e per la superficie interna della membrana cellulare è sempre negativo. È consuetudine misurare il potenziale di riposo sulla superficie interna della membrana, perché la composizione ionica del citoplasma cellulare è più stabile del fluido extracellulare. Il valore del potenziale di riposo è relativamente costante per ogni tipo di cellula. Per le cellule muscolari striate, varia da –50 a –90 mV e per le cellule nervose da –50 a –80 mV.
Le ragioni per l'emergere del potenziale di riposo sono diversa concentrazione di cationi e anioni anche fuori e dentro la gabbia permeabilità selettiva per loro la membrana cellulare. Il citoplasma di una cellula nervosa e muscolare a riposo contiene circa 30-50 volte più cationi di potassio, 5-15 volte meno cationi di sodio e 10-50 volte meno anioni di cloro rispetto al fluido extracellulare.
A riposo, quasi tutti i canali del sodio della membrana cellulare sono chiusi e la maggior parte dei canali del potassio è aperta. Ogni volta che gli ioni di potassio colpiscono il canale aperto, passano attraverso la membrana. Poiché ci sono molti più ioni potassio all'interno della cellula, la forza osmotica li spinge fuori dalla cellula. I cationi di potassio rilasciati aumentano la carica positiva sulla superficie esterna della membrana cellulare. Come risultato del rilascio di ioni potassio dalla cellula, la loro concentrazione all'interno e all'esterno della cellula dovrebbe presto equalizzarsi. Tuttavia, ciò è impedito dalla forza elettrica di repulsione degli ioni potassio positivi dalla superficie esterna della membrana caricata positivamente.
Più diventa il valore della carica positiva sulla superficie esterna della membrana, più è difficile per gli ioni potassio passare dal citoplasma attraverso la membrana. Gli ioni di potassio lasceranno la cellula fino a quando la forza della repulsione elettrica diventa uguale alla forza della pressione osmotica K +. A questo livello di potenziale sulla membrana, l'ingresso e l'uscita degli ioni potassio dalla cellula sono in equilibrio, quindi la carica elettrica sulla membrana in questo momento è chiamata potenziale di equilibrio del potassio... Per i neuroni, è uguale a –80 a –90 mV.
Poiché in una cellula a riposo quasi tutti i canali del sodio della membrana sono chiusi, gli ioni Na + entrano nella cellula lungo un gradiente di concentrazione in quantità insignificante. Solo in minima parte compensano la perdita di carica positiva da parte dell'ambiente interno della cellula causata dal rilascio di ioni potassio, ma non possono sostanzialmente compensare questa perdita. Pertanto, la penetrazione degli ioni sodio nella cellula (perdita) porta solo ad una leggera diminuzione del potenziale di membrana, per cui il potenziale di membrana a riposo ha un valore leggermente inferiore rispetto al potenziale di equilibrio del potassio.
Pertanto, i cationi di potassio che lasciano la cellula insieme a un eccesso di cationi di sodio nel fluido extracellulare creano un potenziale positivo sulla superficie esterna della membrana di una cellula a riposo.
A riposo, la membrana plasmatica della cellula è ben permeabile agli anioni cloro. Gli anioni di cloro, che sono più presenti nel fluido extracellulare, diffondono nella cellula e portano con sé una carica negativa. Non si verifica una completa equalizzazione della concentrazione di ioni cloro all'esterno e all'interno della cella, perché ciò è impedito dalla forza di reciproca repulsione elettrica delle stesse cariche. Creato potenziale di equilibrio del cloro, in cui l'ingresso degli ioni cloro nella cella e la loro uscita da essa sono in equilibrio.
La membrana cellulare è praticamente impermeabile ai grandi anioni degli acidi organici. Pertanto, rimangono nel citoplasma e, insieme agli anioni cloro in ingresso, forniscono un potenziale negativo sulla superficie interna della membrana di una cellula nervosa a riposo.
Il valore più importante del potenziale di membrana a riposo è che crea un campo elettrico che agisce sulle macromolecole di membrana e conferisce ai loro gruppi carichi una certa posizione nello spazio. È particolarmente importante che questo campo elettrico determini lo stato chiuso delle porte di attivazione dei canali del sodio e lo stato aperto delle loro porte di inattivazione (Fig. 61, A). Ciò garantisce lo stato di riposo della cellula e la sua disponibilità all'eccitazione. Anche una diminuzione relativamente piccola del potenziale di membrana a riposo apre le "porte" di attivazione dei canali del sodio, che porta la cellula fuori dallo stato dormiente e dà origine all'eccitazione.
Tutte le cellule viventi hanno la capacità, sotto l'influenza di stimoli, di passare da uno stato di riposo fisiologico a uno stato di attività o eccitazione.
Eccitazioneè un complesso di alterazioni elettriche, chimiche e funzionali attive nei tessuti eccitabili (nervoso, muscolare o ghiandolare), con cui il tessuto risponde alle influenze esterne. Un ruolo importante nell'eccitazione è svolto dai processi elettrici che assicurano la conduzione dell'eccitazione lungo le fibre nervose e portano i tessuti in uno stato attivo (di lavoro).
Potenziale di membrana
Le cellule viventi hanno una proprietà importante: la superficie interna della cellula è sempre caricata negativamente rispetto al suo lato esterno. Esiste una differenza di potenziale tra la superficie esterna della cellula, caricata elettropositivamente rispetto al protoplasma, e il lato interno della membrana cellulare, che varia da 60 a 70 mV. Secondo PG Kostiuk (2001), in una cellula nervosa questa differenza varia da 30 a 70 mV. La differenza di potenziale tra i lati esterno e interno della membrana cellulare è chiamata potenziale di membrana, o potenziale di riposo(fig. 2.1).
Il potenziale di membrana a riposo è presente sulla membrana finché la cellula è viva e scompare con la morte della cellula. L. Galvani nel 1794 dimostrò che se un nervo o un muscolo viene danneggiato facendo una sezione trasversale e applicando elettrodi collegati a un galvanometro alla parte danneggiata e al luogo del danno, allora il galvanometro mostrerà una corrente che scorre sempre da la parte non danneggiata del tessuto al sito dell'incisione ... Ha chiamato questo flusso la corrente di riposo. Per la loro essenza fisiologica, la corrente di riposo e il potenziale di membrana a riposo sono la stessa cosa. La differenza di potenziale misurata in questo esperimento è di 30-50 mV, poiché quando il tessuto è danneggiato, parte della corrente viene deviata nello spazio intercellulare e nel fluido che circonda la struttura in esame. La differenza di potenziale può essere calcolata utilizzando la formula di Nernst:
dove R è la costante dei gas, T è la temperatura assoluta, F è il numero di Faraday, [K] ext. e [K] cuccetta. - la concentrazione di potassio all'interno e all'esterno della cellula.

Riso. 2.1.
La ragione dell'emergere del potenziale di riposo è comune a tutte le cellule. Esiste una distribuzione non uniforme degli ioni (asimmetria ionica) tra il protoplasma della cellula e l'ambiente extracellulare. La composizione del sangue umano in termini di equilibrio salino ricorda la composizione dell'acqua dell'oceano. Anche l'ambiente extracellulare del sistema nervoso centrale contiene molto cloruro di sodio. La composizione ionica del citoplasma delle cellule è più povera. All'interno delle cellule ci sono 8-10 volte meno ioni Na + e 50 volte meno ioni C! " K + nel citoplasma sono anioni organici, in particolare anioni di aspartico, istamina e altri amminoacidi. Tale asimmetria è una violazione della termodinamica equilibrio.Per ripristinarlo, gli ioni di potassio devono gradualmente lasciare la cellula e gli ioni di sodio - tendono ad essa.Tuttavia, questo non va avanti.
Il primo ostacolo all'equalizzazione della differenza nelle concentrazioni di ioni è la membrana plasmatica della cellula. È costituito da un doppio strato di molecole fosfolipidiche, ricoperto dall'interno da uno strato di molecole proteiche, e dall'esterno da uno strato di carboidrati (mucopolisaccaridi). Alcune delle proteine cellulari sono incorporate direttamente nel doppio strato lipidico. Queste sono proteine interne.
Le proteine di membrana di tutte le cellule sono divise in cinque classi: pompe, canali, recettori, enzimi e proteine strutturali. Pompe servono a spostare ioni e molecole contro gradienti di concentrazione utilizzando energia metabolica. Canali proteici, o pori, fornire permeabilità selettiva (diffusione) attraverso la membrana degli ioni e delle molecole ad essi corrispondenti in termini di dimensioni. Proteine del recettore possedendo un'elevata specificità, riconoscono e legano, legandosi alla membrana, molti tipi di molecole necessarie per l'attività vitale della cellula in un dato momento. Enzimi accelerare il corso delle reazioni chimiche sulla superficie della membrana. proteine strutturali garantire la connessione delle cellule negli organi e il mantenimento della struttura subcellulare.
Tutte queste proteine sono specifiche, ma non strettamente. In determinate condizioni, una particolare proteina può essere contemporaneamente una pompa, un enzima e un recettore. Attraverso i canali della membrana, le molecole d'acqua e gli ioni corrispondenti alla dimensione dei pori entrano ed escono dalla cellula. La permeabilità della membrana per diversi cationi non è la stessa e cambia in diversi stati funzionali del tessuto. A riposo, la membrana è 25 volte più permeabile agli ioni potassio rispetto agli ioni sodio e, dopo l'eccitazione, la permeabilità al sodio è circa 20 volte superiore a quella del potassio. A riposo, uguali concentrazioni di potassio nel citoplasma e sodio nell'ambiente extracellulare dovrebbero fornire una quantità uguale di cariche positive su entrambi i lati della membrana. Ma poiché la permeabilità per gli ioni potassio è 25 volte superiore, allora il potassio, uscendo dalla cellula, rende la sua superficie sempre più caricata positivamente rispetto al lato interno della membrana, attorno alla quale molecole cariche negativamente di aspartico, istamina e altre molecole che sono troppo grandi perché i pori della membrana si accumulino amminoacidi che "rilasciano" potassio all'esterno della cellula, ma "non gli permettono" di andare lontano a causa della sua carica negativa. Le cariche negative si accumulano all'interno della membrana e le cariche positive all'esterno. Sorge una potenziale differenza. La corrente diffusa di ioni sodio nel protoplasma dal fluido extracellulare mantiene questa differenza al livello di 60-70 mV, impedendone l'aumento. La corrente diffusa degli ioni sodio a riposo è 25 volte più debole della controcorrente degli ioni potassio. Gli ioni sodio, penetrando nella cellula, riducono il valore del potenziale di riposo, permettendo di mantenerlo ad un certo livello. Pertanto, il valore del potenziale di riposo delle cellule muscolari e nervose, nonché delle fibre nervose, è determinato dal rapporto tra il numero di ioni potassio carichi positivamente, che si diffondono per unità di tempo dalla cellula verso l'esterno, e ioni sodio carichi positivamente, che si diffondono attraverso la membrana nella direzione opposta. Maggiore è questo rapporto, maggiore è il valore del potenziale di riposo e viceversa.
Il secondo ostacolo che mantiene la differenza di potenziale ad un certo livello è la pompa sodio-potassio (Fig. 2.2). Si chiama sodio-potassio o ionico, poiché rimuove attivamente (pompando) gli ioni di sodio che penetrano in esso dal protoplasma e introduce (pompando) ioni di potassio in esso. La fonte di energia per il funzionamento della pompa ionica è la scissione dell'ATP (adenosina trifosfato), che avviene sotto l'influenza dell'enzima adenosina trifosfatasi, localizzato nella membrana cellulare e attivato dagli stessi ioni, cioè potassio e sodio ( ATP-asi sodio-potassio-dipendente).

Riso. 2.2.
È una grande proteina che è più grande dello spessore della membrana cellulare. La molecola di questa proteina, penetrando a fondo nella membrana, lega principalmente sodio e ATP sul lato interno, potassio e vari inibitori di tipo glicosidico sul lato esterno. In questo caso, si verifica una corrente di membrana. Grazie a questa corrente, viene assicurata una direzione appropriata del trasporto ionico. Il trasferimento di ioni avviene in tre fasi. Innanzitutto, lo ione si combina con una molecola di trasporto per formare un complesso di ioni di trasporto. Questo complesso passa quindi attraverso la membrana o trasferisce la carica attraverso di essa. Infine, lo ione viene liberato dal trasportatore sul lato opposto della membrana. Allo stesso tempo, avviene un processo simile, trasferendo gli ioni nella direzione opposta. Se la pompa esegue il trasferimento di uno ione sodio a uno ione potassio, mantiene semplicemente un gradiente di concentrazione su entrambi i lati della membrana, ma non contribuisce alla creazione del potenziale di membrana. Per dare questo contributo, la pompa ionica deve trasferire sodio e potassio in un rapporto di 3:2, cioè per 2 ioni potassio che entrano nella cellula, deve rimuovere 3 ioni sodio dalla cellula. Quando funziona al carico massimo, ogni pompa è in grado di pompare circa 130 ioni potassio e 200 ioni sodio al secondo attraverso il diaframma. Questa è la velocità massima. In condizioni reali, il funzionamento di ciascuna pompa viene regolato in base alle esigenze della gabbia. La maggior parte dei neuroni ha tra 100 e 200 pompe ioniche per micron quadrato di superficie della membrana. Di conseguenza, la membrana di qualsiasi cellula nervosa contiene 1 milione di pompe ioniche in grado di muovere fino a 200 milioni di ioni sodio al secondo.
Pertanto, il potenziale di membrana (potenziale di riposo) viene creato come risultato di meccanismi sia passivi che attivi. Il grado di partecipazione di determinati meccanismi in cellule diverse non è lo stesso, da cui ne consegue che il potenziale di membrana può essere diverso in strutture diverse. L'attività delle pompe può dipendere dal diametro delle fibre nervose: più sottile è la fibra, maggiore è il rapporto tra la dimensione della superficie e il volume del citoplasma, rispettivamente, e l'attività delle pompe necessarie per mantenere la differenza di ioni le concentrazioni sulla superficie e all'interno della fibra devono essere maggiori. In altre parole, il potenziale di membrana può dipendere dalla struttura del tessuto nervoso, e quindi dal suo scopo funzionale. La polarizzazione elettrica della membrana è la condizione principale per l'eccitabilità della cellula. Questa è la sua costante disponibilità all'azione. Questa è una riserva di energia potenziale della cellula, che può utilizzare nel caso in cui il sistema nervoso abbia bisogno della sua reazione immediata.
text_fields
text_fields
arrow_upward
Potenziale di membrana a riposo (MPP) o potenziale di riposo (PP) è chiamata la differenza di potenziale di una cellula a riposo tra i lati interno ed esterno della membrana. Il lato interno della membrana cellulare è caricato negativamente rispetto al lato esterno. Prendendo zero il potenziale della soluzione esterna, l'MPP si scrive con il segno meno. La grandezza WFP dipende dal tipo di tessuto e varia da -9 a -100 mV. Pertanto, a riposo, la membrana cellulare polarizzato. La diminuzione del valore MPP si chiama depolarizzazione, aumento - iperpolarizzazione, ripristinando il valore originale WFP- Rifpolarizzazione membrane.
Disposizioni di base della teoria dell'origine delle membrane WFP riduci a quanto segue. A riposo, la membrana cellulare è ben permeabile agli ioni K+ (in un certo numero di cellule e agli SG), meno permeabile al Na+ e praticamente impermeabile alle proteine intracellulari e ad altri ioni organici. Gli ioni K + diffondono dalla cellula lungo un gradiente di concentrazione, mentre gli anioni non penetranti rimangono nel citoplasma, fornendo l'apparenza di una potenziale differenza attraverso la membrana.
La differenza di potenziale risultante impedisce l'uscita di K + dalla cellula e, ad un certo valore di esso, si verifica l'equilibrio tra l'uscita di K + lungo il gradiente di concentrazione e l'ingresso di questi cationi lungo il gradiente elettrico sorto. Il potenziale di membrana al quale si raggiunge questo equilibrio è detto potenziale di equilibrioscarlatto. Il suo valore può essere calcolato dall'equazione di Nernst:
dove E a- potenziale di equilibrio per A + ; R- costante del gas; T- temperatura assoluta; F - numero di Faraday; NS- valenza K + (+1), [K n +] - [K + est] - concentrazione esterna e interna K + -
Se passiamo dai logaritmi naturali al decimale e sostituiamo i valori numerici delle costanti nell'equazione, l'equazione assumerà la forma: 
Nei neuroni spinali (Tabella 1.1) E k = -90 mV. Il valore MPP misurato con microelettrodi è notevolmente inferiore - 70 mV.
Tabella 1.1... Concentrazione di alcuni ioni all'interno e all'esterno dei motoneuroni spinali dei mammiferi
| E lui |
Concentrazione |
(mmol / l H 2 O) |
Potenziale di equilibrio (mv) |
|
dentro la cellula |
fuori dalla gabbia |
||
| Na + | 15,0 | 150,0 | |
| K + | 150,0 | 5,5 | |
| l - | 125,0 | ||
|
Potenziale di membrana a riposo = -70 mV |
|||
Se il potenziale di membrana di una cellula è di natura potassica, allora, secondo l'equazione di Nernst, il suo valore dovrebbe diminuire linearmente con una diminuzione del gradiente di concentrazione di questi ioni, ad esempio con un aumento della concentrazione di K + nel liquido extracellulare. Tuttavia, la dipendenza lineare del BMP (Resting Membrane Potential) dal gradiente di concentrazione di K + esiste solo quando la concentrazione di K + nel fluido extracellulare è superiore a 20 mM. A concentrazioni inferiori di K + all'esterno della cellula, la curva della dipendenza di E m dal logaritmo del rapporto tra la concentrazione di potassio all'esterno e all'interno della cellula differisce da quella teorica. È possibile spiegare le deviazioni stabilite della dipendenza sperimentale dell'entità dell'MPP e del gradiente di concentrazione K + calcolato teoricamente usando l'equazione di Nernst, assumendo che l'MPP delle cellule eccitabili sia determinato non solo dal potassio, ma anche dal sodio e potenziali di equilibrio del cloro. Ragionando in modo simile al precedente, puoi scrivere:
I valori dei potenziali di equilibrio di sodio e cloruro per i neuroni spinali (Tabella 1.1) sono rispettivamente pari a +60 e -70 mV. Il valore E Cl è uguale al valore MPP. Ciò indica una distribuzione passiva degli ioni cloro attraverso la membrana secondo gradienti chimici ed elettrici. Per gli ioni sodio, i gradienti chimici ed elettrici sono diretti verso l'interno della cellula.
Il contributo di ciascuno dei potenziali di equilibrio al valore MPP è determinato dal rapporto tra la permeabilità della membrana cellulare per ciascuno di questi ioni. Il calcolo del valore del potenziale di membrana viene effettuato utilizzando l'equazione di Goldman:

E m- potenziale di membrana; R- costante del gas; T- temperatura assoluta; F- Numero di Faraday; RK, P Na e RCl - costanti di permeabilità di membrana per K + Na + e Cl, rispettivamente; [A+ n ], [ K + ext, [ N / A+ n [ N / A + ext], [Сl - n] e [Сl - int] sono le concentrazioni di K +, Na + e Cl all'esterno (n) e all'interno (fuori) della cellula.
Sostituendo in questa equazione la concentrazione di ioni e il valore del MPP ottenuto negli studi sperimentali, si può dimostrare che per l'assone gigante del calamaro dovrebbe esserci il seguente rapporto delle costanti di permeabilità P a: P Na: P C1 = Io: 0,04: 0,45. Ovviamente, poiché la membrana è permeabile agli ioni sodio (P N a =/ 0) e il potenziale di equilibrio per questi ioni ha un segno "più", quindi l'ingresso di quest'ultimo nella cellula lungo gradienti chimici ed elettrici ridurrà l'elettronegatività del citoplasma, cioè aumentare il MPP (Potenziale di Membrana a Riposo).
Con un aumento della concentrazione di ioni potassio nella soluzione esterna superiore a 15 mM, l'MPP aumenta e il rapporto delle costanti di permeabilità cambia verso un eccesso più significativo di »P rispetto a P Na e P C1. P a: P Na: P C1 = 1: 0,025: 0,4. In tali condizioni, l'MPP è determinato quasi esclusivamente dal gradiente di ioni potassio; pertanto, le dipendenze sperimentali e teoriche del valore MPP dal logaritmo del rapporto tra le concentrazioni di potassio all'esterno e all'interno della cellula iniziano a coincidere.
Pertanto, la presenza di una differenza di potenziale stazionaria tra il citoplasma e l'ambiente esterno in una cellula a riposo è dovuta ai gradienti di concentrazione esistenti per K +, Na + e Cl e alla diversa permeabilità della membrana per questi ioni. Il ruolo principale nella generazione di MPP è giocato dalla diffusione di ioni potassio dalla cellula nella soluzione esterna. Insieme a questo, l'MPP è anche determinato dai potenziali di equilibrio del sodio e del cloruro e il contributo di ciascuno di essi è determinato dalla relazione tra le permeabilità della membrana plasmatica cellulare per questi ioni.
Tutti i fattori sopra elencati costituiscono il cosiddetto componente ionico MPP (Potenziale di Riposo di Membrana). Poiché né i potenziali di equilibrio del potassio né del sodio sono uguali all'MPP. la cellula deve assorbire Na+ e perdere K+. La costanza delle concentrazioni di questi ioni nella cellula viene mantenuta grazie al lavoro di Na + K + -ATPasi.
Tuttavia, il ruolo di questa pompa ionica non si limita al mantenimento dei gradienti di sodio e potassio. È noto che la pompa del sodio è elettrogena e durante il suo funzionamento si ha un flusso netto di cariche positive dalla cellula nel fluido extracellulare, che provoca un aumento dell'elettronegatività del citoplasma rispetto al mezzo. L'elettrogenicità della pompa del sodio è stata rivelata in esperimenti sui neuroni giganti del mollusco. L'iniezione elettroforetica di ioni Na+ nel corpo di un singolo neurone ha causato l'iperpolarizzazione della membrana, durante la quale l'MPP era significativamente inferiore al valore del potenziale di equilibrio del potassio. Questa iperpolarizzazione è stata indebolita con una diminuzione della temperatura della soluzione in cui si trovava la cellula ed è stata soppressa dall'inibitore specifico di Na +, K + -ATPasi ouabain.
Da quanto sopra consegue che l'MPP può essere suddiviso in due componenti: "Ionico" e "metabolico". Il primo componente dipende dai gradienti di concentrazione degli ioni e dalla loro permeabilità di membrana. Il secondo, "metabolico", è dovuto al trasporto attivo di sodio e potassio ed ha un duplice effetto su MPP. Da un lato, una pompa del sodio mantiene i gradienti di concentrazione tra il citoplasma e l'ambiente. D'altra parte, essendo elettrogena, la pompa del sodio ha un effetto diretto sull'MPP. Il suo contributo al valore MPP dipende dalla densità della corrente di "pompaggio" (corrente per unità di superficie della membrana cellulare) e dalla resistenza della membrana.
Potenziale d'azione di membrana
text_fields
text_fields
arrow_upward
Se l'irritazione viene applicata a un nervo o un muscolo al di sopra della soglia di eccitazione, allora l'MPP del nervo o del muscolo diminuirà rapidamente e per un breve periodo di tempo (millisecondi) la membrana verrà ricaricata: il suo lato interno si caricherà positivamente rispetto a quello esterno. esso una variazione a breve termine dell'MPP che si verifica quando la cella è eccitata, che sullo schermo dell'oscilloscopio ha la forma di un unico picco, è chiamata potenziale d'azione di membrana (MTD).
La IVD nei tessuti nervosi e muscolari si verifica quando il valore assoluto di MPP (depolarizzazione di membrana) scende a un certo valore critico, chiamato soglia di generazione MTD. Nelle fibre nervose giganti del calamaro, l'IVD è - 60 mV. Quando la membrana è depolarizzata a -45 mV (la soglia di generazione dell'IVD), appare l'IVD (Fig. 1.15).
Riso. 1.15 Potenziale d'azione di una fibra nervosa (A) e variazioni della conduttività di membrana per gli ioni sodio e potassio (B).Durante l'insorgenza della IVD nell'assone di calamaro, la resistenza di membrana diminuisce di 25 volte, da 1000 a 40 Ohm.cm 2, mentre la capacità elettrica non cambia. Questa diminuzione della resistenza della membrana è dovuta ad un aumento della permeabilità ionica della membrana all'eccitazione.
In termini di ampiezza (100-120 mV), l'MTD (Membrane Action Potential) è 20-50 mV più alto dell'MPP (Resting Membrane Potential). In altre parole, il lato interno della membrana per un breve periodo si carica positivamente rispetto all'esterno, - "overshoot" o inversione di carica.
Dall'equazione di Goldmann segue che solo un aumento della permeabilità della membrana per gli ioni sodio può portare a tali cambiamenti nel potenziale della membrana. Il valore di E k è sempre inferiore al valore dell'MPP, quindi un aumento della permeabilità della membrana per K + aumenterà il valore assoluto dell'MPP. Il potenziale di equilibrio del sodio ha un segno "più", quindi un forte aumento della permeabilità di membrana per questi cationi porta ad una ricarica della membrana.
Durante l'IVD, la permeabilità della membrana agli ioni sodio aumenta. I calcoli hanno mostrato che se a riposo il rapporto delle costanti di permeabilità della membrana per K +, Na + e SG è 1: 0,04: 0,45, quindi a MPD - P k: P Na: P = 1: 20: 0,45 ... Di conseguenza, in uno stato di eccitazione, la membrana della fibra nervosa non solo perde la sua permeabilità ionica selettiva, ma, al contrario, da permeabile selettivamente a riposo agli ioni potassio, diventa selettivamente permeabile agli ioni sodio. Un aumento della permeabilità al sodio della membrana è associato all'apertura dei canali del sodio voltaggio-dipendenti.
Il meccanismo che prevede l'apertura e la chiusura dei canali ionici si chiama porta del canale.È consuetudine distinguere Attivazione(m) e inattivare(h) cancello. Il canale ionico può trovarsi in tre stati principali: chiuso (m-gate è chiuso; h-aperto), aperto (m- e h-gate sono aperti) e inattivato (m-gate è aperto, h-gate è chiuso) ( Figura 1.16).
Riso. 1.16 Schema della posizione delle porte di attivazione (m) e di disattivazione (h) dei canali del sodio corrispondenti agli stati chiuso (riposo, A), aperto (attivazione, B) e inattivato (C).
La depolarizzazione della membrana, causata da uno stimolo irritante, ad esempio una corrente elettrica, apre la m-gate dei canali del sodio (transizione dallo stato A allo stato B) e fornisce un flusso verso l'interno di cariche positive - ioni sodio. Ciò porta a un'ulteriore depolarizzazione della membrana, che, a sua volta, aumenta il numero di canali del sodio aperti e, quindi, aumenta la permeabilità al sodio della membrana. Si ha una depolarizzazione "rigenerativa" della membrana, per cui il potenziale del lato interno della membrana tende a raggiungere il valore del potenziale di equilibrio del sodio.
Il motivo della cessazione della crescita dell'IVD (Membrane Action Potential) e della ripolarizzazione della membrana cellulare è:
un) Aumento della depolarizzazione della membrana, ad es. quando E m - »E Na, a seguito del quale il gradiente elettrochimico per gli ioni sodio diminuisce, pari a E m -> E Na. In altre parole, la forza che "spinge" il sodio nella cellula diminuisce;
B) La depolarizzazione della membrana dà origine al processo di inattivazione dei canali del sodio (chiusura dell'h-gate; stato del canale B), che inibisce la crescita della permeabilità al sodio della membrana e porta alla sua diminuzione;
v) La depolarizzazione della membrana ne aumenta la permeabilità agli ioni potassio. La corrente di potassio in uscita tende a spostare il potenziale di membrana verso il potenziale di equilibrio di potassio.
Una diminuzione del potenziale elettrochimico per gli ioni sodio e l'inattivazione dei canali del sodio riduce la quantità di corrente di sodio in ingresso. Ad un certo momento, il valore della corrente di sodio in entrata viene confrontato con l'aumento della corrente in uscita: la crescita dell'MPD si interrompe. Quando la corrente totale in uscita supera quella in entrata, inizia la ripolarizzazione della membrana, che ha anche un carattere rigenerativo. La ripolarizzazione incipiente porta alla chiusura della porta di attivazione (m), che diminuisce la permeabilità al sodio della membrana, accelera la ripolarizzazione e quest'ultima aumenta il numero di canali chiusi, ecc.
La fase di ripolarizzazione IVD in alcune cellule (ad esempio nei cardiomiociti e in alcune cellule muscolari lisce) può rallentare, formando altopiano AP, a causa di complesse variazioni nel tempo delle correnti entranti e uscenti attraverso la membrana. L'iperpolarizzazione e/o la depolarizzazione della membrana possono verificarsi nell'effetto collaterale dell'IVD. Questi sono i cosiddetti potenziali di traccia. L'iperpolarizzazione della traccia ha una duplice natura: ionico e metabolicofucina. Il primo è legato al fatto che la permeabilità al potassio nella fibra nervosa della membrana rimane per qualche tempo (decine e anche centinaia di millisecondi) aumentata dopo la generazione di IVD e sposta il potenziale di membrana verso il potenziale di equilibrio del potassio. L'iperpolarizzazione della traccia dopo stimolazione ritmica delle cellule è associata principalmente all'attivazione della pompa del sodio elettrogenica, a causa dell'accumulo di ioni sodio nella cellula.
Il motivo della depolarizzazione che si sviluppa dopo la generazione dell'IVD (Membrane Action Potential) è l'accumulo di ioni potassio sulla superficie esterna della membrana. Quest'ultimo, come segue dall'equazione di Goldmann, porta ad un aumento del MPP (Resting Membrane Potential).
Un'importante proprietà della fibra nervosa è associata all'inattivazione dei canali del sodio, chiamatarefrattarietà .
Durante assolutoferoce periodo refrattario la fibra nervosa perde completamente la sua capacità di essere eccitata dall'azione di un irritante di qualsiasi forza.
Parente refrattarietà, dopo quello assoluto, è caratterizzato da una soglia più alta di IVD (Membrane Action Potential).
Il concetto di processi di membrana che si verificano durante l'eccitazione di una fibra nervosa serve come base per comprendere il fenomeno struttura ricettiva. Al centro dell'accomodamento dei tessuti con una piccola pendenza dell'aumento della corrente irritante c'è un aumento della soglia di eccitazione, che è prima della lenta depolarizzazione della membrana. L'aumento della soglia di eccitazione è quasi interamente determinato dall'inattivazione dei canali del sodio. Il ruolo dell'aumento della permeabilità al potassio della membrana nello sviluppo dell'accomodazione è che porta a un calo della resistenza della membrana. A causa della diminuzione della resistenza, la velocità di depolarizzazione della membrana diventa ancora più lenta. Il tasso di accomodazione è maggiore, maggiore è il numero di canali del sodio a potenziale di riposo in uno stato inattivato, maggiore è il tasso di sviluppo dell'inattivazione e maggiore è la permeabilità al potassio della membrana.
Condurre l'eccitazione
text_fields
text_fields
arrow_upward
La conduzione dell'eccitazione lungo la fibra nervosa viene effettuata a causa di correnti locali tra le sezioni eccitate e a riposo della membrana. La sequenza degli eventi in questo caso è presentata come segue.
Quando si applica una stimolazione puntiforme a una fibra nervosa, nella sezione corrispondente della membrana si genera un potenziale d'azione. Il lato interno della membrana in un dato punto risulta essere caricato positivamente rispetto a quello vicino, a riposo. Una corrente nasce tra punti della fibra che hanno potenziali diversi (corrente locale), diretto da eccitato (segno (+) sul lato interno della membrana) a non eccitato (segno (-) sul lato interno della membrana) alla sezione della fibra. Questa corrente ha un effetto depolarizzante sulla membrana della fibra nell'area di riposo, e quando il livello critico di depolarizzazione della membrana viene raggiunto in quest'area, si verifica un IVD (Membrane Action Potential). Questo processo si diffonde in sequenza a tutte le parti della fibra nervosa.
In alcune cellule (neuroni, muscolatura liscia), l'IVD non è di natura sodica, ma è dovuta all'ingresso di ioni Ca 2+ attraverso canali del calcio voltaggio-dipendenti. Nei cardiomiociti, la generazione IVD è associata alle correnti di sodio e sodio-calcio in ingresso.